
SOMOS QUIMICA
Somos una gran familia cuya raíz es la Química. Lo más asombroso es que todos los seres vivos que habitamos en la Tierra descendemos de un antepasado común. Los animales, los hongos, las plantas, las algas, los microorganismos que nos rodean y se reproducen dentro de nosotros... somos realmente parientes. Formamos parte de una misma familia que lleva más de 3.500 millones de años (Ma) evolucionando y diversificándose. Un árbol cuyas profundas raíces se hunden en la Química que existía en un planeta rocoso situado en la zona de habitabilidad en torno al Sol.
DE DÓNDE VENIMOS
Los cosmólogos explican que hace unos 13.770 Ma una fluctuación cuántica en el vacío, en la nada primigenia, lo originó todo : el tiempo y el espacio, la materia y la energía. Sin entrar a comentar los trascendentales procesos que ocurrieron durante los primeros instantes tras el Big Bang , se ha calculado que 3 minutos después de esa singularidad la temperatura y densidad del Universo habían disminuido lo suficiente como para que se iniciaran las reacciones de «nucleosíntesis primordial». Así, a los 20 minutos de su origen, un 75 % de la masa del Cosmos formaba núcleos de hidrógeno (H), casi el 25 % restante eran núcleos de helio (He), y existían también trazas de sus isótopos, de núcleos de litio (Li) y de berilio (Be). Unos 380.000 años más tarde, el progresivo enfriamiento permitió que los electrones comenzaran a interaccionar electrostáticamente con los núcleos, formando átomos. En ese proceso, llamado «recombinación», el Universo se volvió transparente y la radiación pudo viajar sin obstáculos. Así se produjo el fenómeno más poético de la historia del Cosmos: el origen de la luz.
A partir de las irregularidades existentes en la distribución inicial de materia, hace algo más de 13.300 Ma se formaron las primeras estrellas, compuestas por H y He. En las siguientes generaciones que se fueron sucediendo, las reacciones de «nucleosíntesis estelar» originaron progresivamente todos los elementos de la tabla periódica, hasta llegar al hierro (Fe). Para estrellas de entre 0,5 y 9 masas solares, dichos elementos se acabarían dispersando por el espacio en la fase de su evolución conocida como «gigante roja», originando nebulosas: regiones heterogéneas de gas y polvo con diferente contenido en elementos distintos del H y el He (que en astrofísica se llaman genéricamente «metales»). Entre los elementos sintetizados estaban los cinco que, junto al H, son fundamentales para formar las moléculas de la vida tal como la conocemos: carbono (C), nitrógeno (N), oxígeno (O), fósforo (P) y azufre (S): el famoso grupo «CHONPS», del que hablaremos. Sin duda, como decía Carl Sagan, "somos polvo de estrellas".
Por su parte, de forma simplificada podemos decir que en la explosión de las supernovas resultantes de la evolución de las estrellas más masivas se formaron los elementos de número atómico mayor que el Fe, mientras que los más pesados que el oro (Au) fueron originados principalmente en colisiones de estrellas de neutrones. En las nebulosas, los granos de polvo tienen dimensiones micrométricas. Sus núcleos, formados por silicatos (fundamentalmente compuestos por O y silicio, Si) y grafito (una de las formas estables del C), están recubiertos por hielos de agua (H2 O) y también de otras moléculas como monóxido de carbono (CO), dióxido de carbono (CO2 ) o metanol (CH3OH). El calentamiento de dichos gránulos producido por la actividad estelar, así como la incidencia sobre ellos de radiación cósmica, aportan la energía necesaria para que estas pequeñas moléculas (y los radicales derivados de ellas, con uno o más electrones desapareados y por lo general muy inestables) adquieran movilidad y reaccionen entre sí, dando lugar a compuestos de mayor tamaño. Con ello, la Química puede comenzar a desplegar todo su potencial.
En el medio interestelar y circumestelar, los investigadores del campo de la Astroquímica ya han detectado más de 200 moléculas con un número de átomos típicamente entre 4 y 12, junto a hidrocarburos lineales y cíclicos e incluso fullerenos de 60 y 70 átomos de C. Entre estos compuestos, algunos como el formaldehído (HCHO), glicolaldehído (CH2 OH–CHO, el azúcar más sencillo que existe), cianuro de hidrógeno (HCN), cianoacetileno (HC≡C–CN), cianamida (NH2 CN), formamida (CHO–NH2 ), urea (NH2 –CO–NH2 ) o monóxido de fósforo (la molécula P–O) son especialmente interesantes, ya que pueden funcionar como precursores de las reacciones de Química prebiótica.
En paralelo al progresivo enriquecimiento de su diversidad química, en las nubes moleculares aparecen algunas regiones más densas que el resto, en las cuales se forman núcleos de condensación de tamaño creciente. Cuando la masa de estos núcleos es suficientemente grande se inicia el colapso gravitacional que conduce a la formación de nuevas estrellas. En torno a ellas, el material remanente origina discos protoplanetarios, como ocurrió en las inmediaciones del Sol (una estrella que se considera de tercera generación) hace aproximadamente 4.570 Ma. Dentro de los discos, al irse agregando los granos de polvo que he descrito se forman glóbulos milimétricos, éstos dan lugar a rocas... y ellas a cuerpos llamados «planetesimales» cada vez más grandes.
Continuando el proceso de interacción gravitatoria, la «acreción» progresiva de planetesimales acaba originando planetas de diferentes tamaños y composiciones químicas. En nuestro caso, en la región más interior del Sistema Solar se formaron planetas pequeños y rocosos, mientras que a mayores distancias del Sol se originaron los gigantes de gas, con Júpiter a la cabeza. El tercer planeta que se formó alrededor del Sol, y que había ido aumentando su masa mediante ese proceso de acreción, recibió hace unos 4.540 Ma el impacto de un gran protoplaneta del tamaño de Marte, denominado Tea (como la titánide madre de Selene, diosa de la Luna). Esa enorme colisión extrajo gran cantidad de material de la corteza y el manto de la Tierra, que quedó orbitando nuestro planeta como un disco de escombros. Las interacciones gravitatorias entre esos fragmentos irían generando cuerpos progresivamente mayores, hasta formarse la Luna. Así, el sistema Tierra-Luna se estabilizó hace unos 4.510 Ma: desde entonces Gea y Selene están gravitacionalmente unidas, influyéndose mutuamente y mirándose día y noche... aunque cada año 3,8 centímetros más lejos.
LA TIERRA PRIMITIVA
Durante más de 100 Ma, la Tierra y la Luna estuvieron sometidas a un intenso bombardeo por los fragmentos de roca y hielo que seguían presentes en un Sistema Solar aún en construcción. Además, nuestro planeta mantenía un elevado calor interno debido a la desintegración radiactiva de algunos de los materiales que lo habían originado. El efecto combinado de ambos procesos generaba una temperatura superficial que hacía imposible la existencia de agua líquida en ella, por lo que tanto el agua contenida en los planetesimales originales como la aportada masivamente por los meteoritos y núcleos cometarios se iba evaporando y formaba gruesas capas de nubes alrededor de la Tierra.
La progresiva disminución de la intensidad del bombardeo meteorítico hizo que la superficie y la atmósfera terrestres se fueran enfriando. Se estima que hace unos 4.400 Ma ya se había formado una corteza sólida en la Tierra, al cristalizar el océano de lava fundida presente durante el comienzo del eón Hadeico (un nombre muy adecuado, sin duda). La enorme cantidad de vapor de agua existente en las nubes fue condensándose, lo que produjo lluvias torrenciales durante millones de años: se formaron ríos, torrentes y cataratas, el agua fue disolviendo y arrastrando muchas de las moléculas presentes en las superficies de las rocas, y las zonas más bajas de la corteza acabaron acogiendo un inmenso océano. La Tierra comenzaba a ser un planeta habitable. Y todo indica que en esa época también lo eran Venus y Marte, nuestros vecinos en el Sistema Solar, aunque su historia fue muy diferente.
Existe una notable discrepancia sobre las características físico-químicas de esa hidrosfera terrestre primitiva, que comenzó a interaccionar con la litosfera y con la atmósfera: su temperatura media podría haber sido de entre 40 °C y 80 °C, su salinidad se estima entre 1,5 y 2,5 veces mayor que la de los océanos actuales, y su nivel de acidez o basicidad podría corresponder a valores de pH entre 4 y 9 (recordemos, sobre una escala que va de 0 a 14, con la neutralidad en el pH 7). En cualquier caso, hemos de pensar que el planeta tendría entornos geológicos cada vez más diferenciados, en los que se podrían producir diferentes reacciones químicas, tanto en medio líquido como en su interfase con las rocas.
La atmósfera de aquella Tierra primitiva (una vez perdida en el espacio la envuelta inicial de hidrógeno molecular, H2 , y con el agua ya condensada desde su fase vapor a líquida) era el resultado de la desgasificación de los magmas de la corteza y de las reacciones promovidas en los gases por la radiación solar, que en el rango del ultravioleta era de 5 a 10 veces más intensa que la actual. Así, se estima que hace entre 4.400 y 4.000 Ma la atmósfera terrestre estaría formada principalmente por nitrógeno molecular (N2 ) y dióxido de carbono, con presencia de otros gases como vapor de agua y monóxido de carbono, o en menor proporción sulfuro de hidrógeno (H2 S), metano (CH4 ) e hidrógeno molecular. Durante ese período, la frecuencia de impactos meteoríticos y cometarios que recibió nuestro planeta disminuyó notablemente, por lo que la mayor estabilidad de las aguas y tierras emergidas permitió a la Química explorar diferentes escenarios y probar los sistemas catalíticos disponibles. Así pudieron irse sintetizando moléculas orgánicas progresivamente más complejas, a partir de los precursores que (como hemos visto) formaban parte de los planetesimales originales, junto a los compuestos aportados por los mensajeros del espacio que nos habían llegado incesantemente durante la época anterior.
Por tanto, la Tierra era un laboratorio de dimensiones planetarias que ya contenía los tres ingredientes imprescindibles para la aparición de los seres vivos: agua en estado líquido, moléculas orgánicas y fuentes de energía. Tal vez los primeros experimentos químicos que dieron lugar a la vida tuvieron éxito en esta época tan temprana. Esa posibilidad se ha visto apoyada por el descubrimiento de gránulos de grafito en rocas de entre 4.100 y 3.950 Ma de antigüedad, en los que la relación de isótopos del carbono muestra un desbalance hacia el más ligero, el 12 C o carbono-12: dado que los seres vivos que fijan C a partir del CO2 atmosférico (llamados autótrofos) tienen preferencia por este isótopo frente al 13 C y al 14 C , tal hallazgo podría indicar que ya había vida en esta etapa tan temprana. Sin embargo, también se conocen sistemas abióticos (es decir, no biológicos) que podrían explicar dicha relación isotópica, por lo que realmente resulta imposible determinar si la vida terrestre es tan antigua.

Pero, más allá de nuestro propio planeta, el gigante Júpiter seguía condicionando la dinámica de los cuerpos que formaban el Sistema Solar. Los datos disponibles y los modelos computacionales indican que hace unos 4.100 Ma su órbita sufrió una importante fluctuación, y como resultado «empujó» a un gran número de asteroides hacia el interior del sistema, alcanzando de pleno las órbitas de Marte y la Tierra. Así, nuestro planeta volvió a sufrir un intenso bombardeo meteorítico, que para diferenciarlo del inicial suele denominarse «tardío» (o «Late Heavy Bombardment», LHB), desde hace aproximadamente 4.000 Ma hasta hace unos 3.850 Ma. Si la vida ya se había iniciado en la Tierra, quizá fue eliminada por el efecto «esterilizador» de las temperaturas y presiones asociadas a tantos impactos.
Es imposible saberlo. Pero sí está claro que el LHB aportó un repertorio aún mayor de moléculas orgánicas, que desde su llegada se irían combinando con las previamente formadas en nuestro planeta. Cuando tal proceso terminó, la Tierra adquirió su tamaño definitivo, con un diámetro medio de aproximadamente 12.740 km, y su órbita se estabilizó a unos 150 millones de kilómetros del Sol (este valor, correspondiente al semieje mayor de la órbita elíptica terrestre, se define como una «unidad astronómica», ua). Comenzaba una época más tranquila en la historia de un planeta que pronto mostraría toda su creatividad.
El lienzo estaba listo, la paleta rebosaba de colores y la Química había ido preparando sus pinceles: ya podían comenzar a darse los primeros trazos hacia la Bioquímica, en lo que se acabaría convirtiendo en el retrato de un ser vivo. Pero, sin un pintor ni un plan de trabajo establecido, la naturaleza probaría numerosas combinaciones de tonos, pinceladas con distintas formas y texturas, se confundiría, lo emborronaría todo, encontraría soluciones valiosas sin haberlo previsto, superpondría capas una y otra vez. No había ningún objetivo. Y tampoco prisa. Pero sobraban energía, colores y disolvente para seguir pintando sin parar. Al cabo del tiempo, entre los muchos lienzos inacabados que se amontonaban en el estudio, podríamos reconocernos en al menos uno de los cuadros que allí quedaron expuestos.
EN BUSCA DEL ORIGEN DE LA VIDA
El origen (u orígenes) de la vida es una de las cuestiones más relevantes que nos podemos plantear desde la ciencia. No contamos con evidencias directas sobre ello, y aunque las leyes de la física y la Química nos ayudan a diferenciar entre lo que pudo y no pudo ocurrir en el amanecer de la biología, las dudas superan ampliamente a las certezas. De hecho, siendo honestos, es complicado saber cómo empezó la vida. Eso sí, cada vez vamos acotando mejor el repertorio de preguntas clave que salen a nuestro paso en este fascinante camino: ¿cuándo ocurrió?, ¿cómo se produjo esa transición entre lo no vivo y lo vivo?, ¿cuál fue el papel del azar y la necesidad en ese proceso?, ¿hubo un origen único o pudo haber varios orígenes?, ¿la vida comenzó en la Tierra, o su origen fue extraterrestre? Incluso, durante las dos últimas décadas nos estamos planteando algo aún más inquietante: ¿seremos capaces de reproducir un posible origen de la vida en los laboratorios, lo que supondría crear un ser vivo a partir de sus constituyentes químicos?
Las primeras aproximaciones a la naturaleza de la vida y su posible origen fueron planteadas por los filósofos presocráticos, cuya hipótesis de la «generación espontánea» sería posteriormente apoyada por Aristóteles y su escuela desde el siglo IV antes de nuestra era. Según esta intuitiva idea, los seres vivos surgen espontáneamente a partir de la materia no viva en determinadas condiciones: por ejemplo, los pulgones parecían nacer del rocío que se deposita sobre las plantas de madrugada, o las moscas y gusanos aparentemente se originaban desde la carne en descomposición. A pesar de que esta hipótesis carecía de cualquier base científica, los intentos de refutarla no comenzaron hasta el siglo XVII . Y fue el microbiólogo Louis Pasteur quien, en una serie de elegantes experimentos realizados entre 1859 y 1864, demostraría, en sus propias palabras, que «la generación espontánea es una quimera». Pero, si toda vida proviene de otra vida, quedaba flotando en el aire una pregunta fundamental: ¿cómo pudo surgir el primer ser vivo?
En aquella época dorada de la ciencia, un ilustre contemporáneo de Pasteur fue el naturalista Charles R. Darwin, y es a él a quien debemos las primeras reflexiones genuinamente científicas sobre el origen de la vida. En su libro más justamente famoso, "El origen de las especies", demostró con gran nivel de detalle y profundidad algo que Alfred R. Wallace también había descubierto en paralelo: la selección natural es el motor de la evolución biológica. Pero quizá lo más original de Darwin fue incluir en el sugerente párrafo final de esta obra una frase que pasaría a la historia: «Hay grandeza en esta concepción de que la vida, con sus diferentes fuerzas, ha sido alentada inicialmente en un corto número de formas o en una sola». Así lo escribió en su primera edición, publicada el 24 de noviembre de 1859 y agotada ese mismo día, aunque para la segunda se vio obligado (entre otros, por su esposa Emma) a introducir la expresión «por el Creador» a continuación de la palabra «inicialmente». En cualquier caso, sin censura o con ella, proponer que toda la biodiversidad que observamos proviene de un origen común era revolucionario, e inauguraba una línea de pensamiento que hoy en día conocemos como «del presente hacia el pasado» o topdown : estudiando comparativamente los seres vivos actuales (y los fósiles de formas de vida extintas) podemos encontrar pistas sobre su origen común.
Ese es el postulado que nos ha ido permitiendo construir árboles evolutivos a partir de datos morfológicos, bioquímicos o genéticos de los seres vivos. Poco después, el propio Darwin fue quien sentó las bases de la aproximación complementaria para llegar al origen de la vida, en una inspirada carta que envió a su amigo, el botánico Joseph D. Hooker, el 1 de febrero de 1871. En ella le planteaba que la vida podría haberse originado en «una pequeña charca de agua templada que contuviera todo tipo de sales de fósforo y amonio, luz, calor, electricidad, etc., en la cual un compuesto proteico se formara químicamente, quedando listo para sufrir cambios aún más complejos». Hoy denominamos a esta línea de investigación «del pasado al presente» o bottom-up , y consiste en proponer y realizar reacciones químicas viables en las condiciones de la Tierra primitiva que puedan llevar a la aparición de los seres vivos.
El campo del origen de la vida apenas avanzó durante el medio siglo siguiente, hasta que dos grandes científicos, de manera independiente, consolidaron la aproximación bottom-up al plantear los primeros modelos sobre la emergencia de los seres vivos desde la Química existente en la infancia de nuestro planeta: el bioquímico Alexander I. Oparin en 1924 (a quien, entre otras muchas ideas inspiradoras, le debemos la certera metáfora de la «sopa primitiva») y el biólogo evolutivo John B. S. Haldane en 1929. Sus modelos, similares en algunos aspectos pero con diferencias notables, elevaban la investigación sobre el origen de la vida a la categoría de disciplina científica.
LA QUÍMICA PREBIÓTICA
Tomando como base los modelos de Oparin y Haldane, el siguiente protagonista fue el químico Stanley L. Miller, quien en 1953 realizó en el laboratorio de Harold C. Urey un experimento cuyos resultados se hicieron rápidamente famosos. Su idea era comprobar si de una mezcla de gases como la supuesta para la atmósfera de la Tierra primitiva (que en esa época se consideraba muy reductora, con compuestos ricos en hidrógeno) podría surgir alguna biomolécula relevante para la vida. Con ese objetivo diseñó y construyó un sistema experimental de vidrio, cerrado y previamente esterilizado, que contenía dos matraces: en uno puso agua y la llevó a ebullición, y en el otro recreó la atmósfera mezclando el vapor de agua con metano, amoníaco (NH3 ) e hidrógeno molecular. Sometió dicha mezcla a descargas eléctricas de 60.000 voltios que simulaban las fuentes de energía presentes en la Tierra primitiva, entre ellas las tormentas, el vulcanismo y los impactos de meteoritos o cometas. Al cabo de unos días pudo comprobar que el interior del matraz de reacción se había teñido de color marrón, lo que sugería la formación de compuestos orgánicos en esas condiciones experimentales.
Tras analizar dicho material, Miller descubrió que su fracción soluble contenía gran número de aminoácidos (entre ellos, tres de los que constituyen las proteínas: glicina, alanina y ácido aspártico), y otras muchas moléculas orgánicas hoy presentes en los seres vivos. En variantes posteriores del experimento se produjeron más aminoácidos de los contenidos en nuestras proteínas, aunque algunos (como arginina, glutamina, triptófano y tirosina) nunca se han obtenido de esta forma y probablemente fueron biosintetizados empleando ya rutas bioquímicas desarrolladas en los seres vivos.
La aproximación experimental de Miller demostraba que algunas de las primeras biomoléculas en el camino hacia la vida se pueden originar de forma abiótica y a partir de moléculas muy sencillas, siempre que se disponga de una fuente de energía. Una prueba a favor de la validez de este tipo de síntesis (en principio, posible tanto en la Tierra como en entornos extraterrestres) llegó con la caída en 1969 del meteorito Murchison, una condrita carbonácea: cuando se analizó su materia orgánica, mostró una composición química muy similar a la obtenida en los experimentos de Miller (el original de 1953 y otros posteriores). Sin embargo, como hemos visto, desde hace medio siglo se propone una atmósfera primitiva mucho menos reductora, y se ha comprobado que experimentos similares al de Miller utilizando esas mezclas de gases producen mucha menor cantidad y variedad de biomoléculas. Por tanto, si nuestra atmósfera inicial era oxidante, quizá parte de los aminoácidos que forman nuestras proteínas nos tuvieron que llegar a bordo de meteoritos y núcleos cometarios. En cualquier caso, este trabajo pionero de Miller demostraba que el origen de la vida es abordable por la ciencia experimental, por lo que las preguntas podían trasladarse desde los despachos y bibliotecas a los laboratorios. Con ello, además, se inauguraba una nueva disciplina científica: la Química prebiótica.
El segundo pionero de la investigación experimental sobre el origen de la vida fue el bioquímico Joan Oró, quien, trabajando en Houston, demostró en 1961 que cinco moléculas de ácido cianhídrico (la molécula HCN que ya conocemos, en disolución acuosa) podían combinarse entre sí para formar adenina, una de las cuatro bases nitrogenadas presentes en los nucleótidos de cuya polimerización surgen los ácidos nucleicos, RNA y DNA. Por tanto, si Miller había propuesto una posible vía de síntesis para los monómeros que forman las proteínas, Oró estaba mostrando cómo pudo surgir uno de los componentes clave de los ladrillos estructurales de los polímeros biológicos que almacenan y transmiten la información genética. Desde entonces, la Química prebiótica es una disciplina en constante desarrollo. Gracias al trabajo de muchos científicos se ha descubierto un buen número de reacciones que, partiendo de las moléculas sencillas y las condiciones que podrían existir en la Tierra primitiva, dan lugar a biomoléculas como aminoácidos, nucleótidos, azúcares o lípidos simples.
Una de las preguntas abiertas en este campo es el origen de la «homoquiralidad», es decir, por qué la vida escogió una de las dos formas estructurales (que llamamos L y D, simétricas entre sí como nuestras manos izquierda y derecha) que pueden presentar los aminoácidos, los azúcares y los nucleótidos: aún no sabemos el motivo, pero todas las proteínas están formadas por aminoácidos L, mientras que el DNA y el RNA contienen nucleótidos D. Así, el tipo de homoquiralidad elegida para cada biopolímero es una de las características que definen la vida terrestre, y permitiría distinguirla de otra con un origen diferente.
La aproximación bottom-up también está mostrando cómo, utilizando superficies minerales u otros catalizadores inorgánicos, se pudieron comenzar a polimerizar los aminoácidos para formar péptidos, y los nucleótidos para originar cadenas cortas de ácidos nucleicos. Durante los últimos años ha irrumpido con fuerza en este campo la denominada «Química prebiótica de sistemas», que plantea escenarios más complejos (y por tanto más realistas) para la aparición de las primeras mezclas heterogéneas de biomoléculas de cuya combinación pudo surgir la vida.
DE LA QUÍMICA A LA BIOLOGÍA
Desde la Química prebiótica hasta la aparición de los primeros seres vivos tuvo que recorrerse un largo y complicado camino, pues para que un sistema pueda reproducirse y evolucionar ha de combinar tres componentes fundamentales: un compartimento, una red metabólica y una molécula con información heredable. Por ello, la pregunta clave que se planteaba desde la época de Miller y Oró era cuál de los tres biopolímeros principales de los seres vivos apareció antes. En todas las células, la información genética está codificada en la secuencia de nucleótidos del DNA, y se expresa en el sentido DNA → RNA → proteína. Pero la replicación del DNA es realizada por una proteína (una de las llamadas «enzimas», que catalizan prácticamente todos los procesos que ocurren en las células), por lo que estamos ante una paradoja similar a la del huevo y la gallina, aunque en el ámbito de la biología molecular: sin DNA no pudieron surgir las proteínas... y viceversa.
Tras varias décadas de investigación, creo que la clave está en la molécula intermediaria entre ambas, el RNA, debido a su versatilidad estructural y funcional. Así, las evidencias experimentales apoyan la hipótesis del «Mundo RNA», según la cual este biopolímero habría sido anterior al DNA y a las proteínas porque podía funcionar simultáneamente como genotipo (archivo de información genética) y como fenotipo (catalizador de reacciones, entre ellas quizá su propia replicación), que son las dos caras de la moneda de la vida. El modelo del Mundo RNA todavía tiene importantes cuestiones que resolver, pero en diferentes laboratorios se realizan experimentos de «evolución in vitro » para tratar de identificar distintos tipos de RNA funcionales, como «aptámeros» (que se unen con gran afinidad y especificidad a sus moléculas diana) y «ribozimas» (que catalizan procesos de forma análoga a como lo hacen las enzimas proteicas). Uno de los objetivos sería llegar a encontrar una molécula de RNA capaz de autorreplicarse, similar a las que podrían haber protagonizado la transición entre la bioquímica y la biología.
En paralelo a esta línea de trabajo, que va en busca de la «replicación primordial», otros científicos defienden el «metabolismo primigenio», manteniendo que antes de la aparición de la primera molécula autorreplicativa fue necesario establecer una red de reacciones químicas auto mantenidas en el interior de un compartimento. Afortunadamente, los avances en Química prebiótica de sistemas han aproximado estas dos líneas de investigación, ya que según comenté, sólo de la combinación exitosa de compartimento, metabolismo y replicación pudieron surgir los primeros seres vivos. Con ello, además, se propone que los escenarios tradicionalmente considerados como alternativos y excluyentes para el origen de la vida (por ejemplo, los pequeños charcos de agua templada en la superficie, las interfases agua/roca o las surgencias hidrotermales submarinas) pudieron en realidad ser complementarios en la Tierra primitiva: cada uno de ellos facilitaría la síntesis de una serie de moléculas o de estrategias bioquímicas que más tarde se combinarían en el camino hacia la biología.
Teniendo todo esto en cuenta, la versión actualizada del modelo del Mundo RNA propone que entre la Química prebiótica y las primeras células hubo una etapa protagonizada por protocélulas denominadas «ribocitos»: en ellas el RNA funcionaría como genoma, y las reacciones metabólicas necesarias para el mantenimiento del sistema serían realizadas por ribozimas y otras moléculas catalíticas (entre ellas péptidos o determinados compuestos orgánicos presentes en la Tierra primitiva). Probablemente en esa época surgieron los primeros virus con genoma de RNA, como moléculas capaces de utilizar o «parasitar» la maquinaria que se estaba optimizando en los ribocitos: así lograrían que su propia secuencia de nucleótidos fuera replicada (produciendo muchos errores o mutaciones) y podrían migrar entre protocélulas, «infectándolas» y repitiendo el proceso.
En los laboratorios, la investigación con virus RNA actuales (formados por un genoma, una cubierta de proteínas y en ocasiones una membrana) y viroides (parásitos más sencillos, sólo encontrados en plantas y que están constituidos por una única molécula de RNA de pequeño tamaño) nos ayuda a entender la dinámica replicativa y las capacidades adaptativas en aquel lejano Mundo RNA. Sin entrar en detalles, cuando en esos ribocitos comenzara a traducirse a proteínas, parte de la información del RNA (lo que implicaría que ya se había establecido el código genético y se habían originado los ribosomas primitivos) pudo darse el paso a un «Mundo RNA-proteínas». Más adelante, algunas proteínas catalíticas comenzarían a copiar la secuencia de nucleótidos del RNA en otro tipo de ácido nucleico, el DNA, cuyos monómeros se podrían haber originado con anterioridad y que resultaría más estable como archivo de información. Surgía así el «Mundo DNA-RNA-proteínas», que desde entonces ha escrito la historia de la vida.
En esa época aparecerían los primeros virus con genoma de DNA que, como en el caso anterior, han seguido siendo compañeros inseparables del mundo celular.
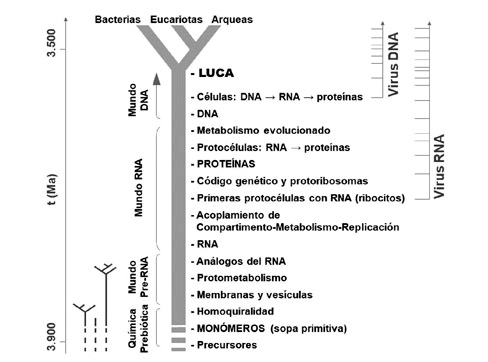
Principales eventos ocurridos durante el origen y la evolución temprana de la vida en la Tierra, a lo largo del tronco del árbol que conecta los precursores químicos con LUCA. Los árboles evolutivos más pequeños muestran la posibilidad de que la vida surgiera más de una vez y se diversificara hasta cierto punto en distintos entornos de nuestro planeta. Se indica también las posibles épocas en las que se originaron los virus con genoma de RNA y de DNA.
MI APELLIDO ES LUCA
En la metáfora del árbol de la vida, una de las más sugerentes que habitualmente utilizamos, la Química prebiótica forma las raíces que encuentran su sustento en las características físico-químicas de la Tierra primitiva. Las biomoléculas de complejidad creciente y las protocélulas que hemos comentado van ocupando posiciones cada vez más altas a lo largo del tronco del árbol, hasta llegar al punto en el que éste empieza a ramificarse. Justo en ese lugar, antes de que comience la diversificación de la vida, es donde estaría el antepasado universal de todos los seres vivos planteado por Darwin al final de "El origen de las especies". Pero ¿realmente existió un ancestro común de toda la biodiversidad que nos rodea? La aproximación top-down hacia el origen de la vida se ha basado en la comparación de las características fenotípicas y genotípicas de buena parte de las especies conocidas.
Desde finales de la década de 1970, la posibilidad de conocer la secuencia de determinados genes en los genomas de numerosos seres vivos permitió establecer una nueva disciplina, denominada «filogenia molecular». Utilizando sus herramientas bioinformáticas es posible traducir distancias genéticas entre dos especies (medidas como el número de mutaciones que diferencian la secuencia del gen comparado) en el tiempo transcurrido desde la divergencia entre ambas. Si se realiza este proceso de forma sistemática para un gran número de especies, trabajo en el que destacó el biofísico y microbiólogo Carl R. Woese, se obtienen árboles filogenéticos que constituyen el punto de encuentro entre la taxonomía y la evolución. De esta forma, durante la década de 1980 se logró demostrar que sí existió el antepasado global predicho por Darwin casi 130 años antes. Se denominó «progenote» o LUCA (por las iniciales en inglés de Last Universal Common Ancestor o «último ancestro común universal») a esa especie celular de la que derivamos todos los seres vivos conocidos. Quizá nuestro nombre no sea Luca, pero nuestro último apellido sí lo es.
Evidentemente, pensando en ese inmenso laboratorio que era la Tierra primitiva, nada impide que pudieran producirse diferentes orígenes de la vida en distintos entornos, como arbustos y pequeños árboles creciendo en el mismo suelo de nuestra metáfora. Pero LUCA representa el punto culminante del único origen de la vida que resultó suficientemente robusto como para resistir las presiones selectivas que estarían operando, y que logró generar la biodiversidad que ha dominado la Tierra desde entonces.
Cronológicamente, suponemos que LUCA vivió entre hace 3.900 y 3.700 Ma, cuando se iniciaba esa época más tranquila de la Tierra primitiva que he comentado. No se pueden estudiar directamente las características que tuvo LUCA, pero sí sabemos que era una especie unicelular, quizá similar a las actuales bacterias aunque con un genoma (de DNA) muy reducido, y que sus menos de 1.000 genes le permitían realizar todas las funciones celulares básicas. Un tema en discusión es si LUCA era autótrofo (es decir, podía extraer energía de los compuestos reducidos presentes en las rocas o bien de la luz solar, fijando carbono desde el CO2 ) o heterótrofo (su metabolismo utilizaba los compuestos orgánicos disponibles en el medio). De hecho, muchos autores prefieren hablar no de un LUCA, sino de varios «LUCAs», para referirse a la comunidad de entidades celulares, genética y metabólicamente complementarias entre sí, que pudieron convivir en la parte más alta del tronco común del árbol de la vida.
A partir de LUCA (o de LUCAs), la evolución biológica condujo a la progresiva diversificación de todas las formas de vida, tanto extintas (de las que en algunos casos tenemos información por sus fósiles) como actuales. Expresando en lenguaje de hoy lo que Wallace y Darwin descubrieron durante la primera mitad del siglo XIX , podríamos decir que la evolución por selección natural se basa en que los errores inevitablemente producidos durante la replicación de los genomas de los seres vivos (mutaciones y otro tipo de reordenamientos genéticos) originan un cierto grado de diversidad fenotípica en su descendencia. Así, los miembros de la progenie responderán de forma diferente a las presiones selectivas que introduzca el ambiente, y los individuos (o las especies) que mejor se adapten generarán más descendencia. Ese proceso, ciego, imprevisible y maravilloso, es el que nos ha traído hasta aquí. Como el genetista Theodosius Dobzhansky tituló en uno de sus ensayos, «Nada tiene sentido en biología excepto a la luz de la evolución».
AZAR, NECESIDAD... ¿Y PANSPERMIA?
Como acabamos de ver, actualmente pensamos que nuestro planeta comenzó a estar habitado poco después de empezar a ser habitable. Por tanto, el origen de la vida habría sido un proceso muy rápido (a escala geológica), y para algunos autores esto indica que la emergencia de la biología a partir de la Química responde a un cierto determinismo o «necesidad» inherente a la materia, que conduciría a dinámicas autoorganizativas y autorreplicativas una vez que se alcanza un determinado grado de complejidad molecular. En ese caso sería probable que la vida hubiese surgido en numerosos lugares del Universo, y además lo habría hecho siguiendo unos principios generales (no necesariamente unas biomoléculas) comunes. Pero, por el contrario, el origen pudo deberse a una sucesión muy afortunada de eventos de azar, casualidades que quizá sólo se habrían podido producir en nuestro planeta y por tanto estaríamos solos en el Cosmos.
Así, desde que Demócrito planteó que «todo lo que existe en el Universo es fruto del azar y la necesidad», cuando algunos científicos han reflexionado sobre el origen de la vida se han mostrado más partidarios de la necesidad (destacando entre ellos el químico Robert Shapiro, el paleontólogo Stephen Jay Gould o el biólogo celular Christian de Duve), mientras que otros han sido firmes defensores del azar (como argumentó el biólogo Jacques L. Monod en su famoso libro "El azar y la necesidad. Ensayo sobre la filosofía natural de la biología moderna". Entre esas dos posturas extremas, probablemente en el camino hacia la vida se combinaron ambos procesos: a partir de una Química compleja con tendencia a autoorganizarse pudieron existir eventos casuales o no deterministas que condujeron a la aparición del primer sistema capaz de autorreproducirse y evolucionar. Y una vez que entra en escena la reproducción (siempre asociada a su correspondiente tasa de error) los caminos que puede seguir la vida (sin desligarse de una cierta necesidad, ya que la selección actúa en cada momento sobre lo que previamente existe) comenzarían a estar fuertemente condicionados por el azar.
Un proceso totalmente dependiente de las casualidades, con el que además se busca resolver la cuestión del poco tiempo transcurrido hasta que apareció la vida en nuestro planeta, es la hipótesis conocida como «panspermia microbiana» o «litopanspermia». Según esta idea, los primeros seres vivos unicelulares se habrían originado fuera de la Tierra, llegando hasta aquí tras un largo viaje a bordo de meteoritos o de núcleos de cometas. Esta hipótesis es muy antigua, pues ya Anaxágoras había sugerido en el siglo V antes de nuestra era que las «semillas o infinitesimales de la vida» podrían formarse en todos los lugares del Cosmos donde las condiciones fueran favorables. En 1865, el biólogo Hermann Richter retomó aquellas ideas para proponer (sin apoyarse en pruebas experimentales) que los microorganismos están presentes en todo el Universo y pudieron «inocular» la vida en nuestro planeta. El químico Svante A. Arrhenius continuó esta corriente a partir de 1908, manteniendo que la vida se había originado en el espacio exterior y los microorganismos viables (o sus esporas) pudieron ser transportados hasta la Tierra gracias a la acción de la presión de la radiación estelar.
Durante el último medio siglo, dicha hipótesis ha sido llevada hasta su extremo por los astrónomos Fred Hoyle y Chandra Wickramasinghe, afirmando incluso que cada vez que nuestra especie sufre una epidemia o pandemia es porque el patógeno correspondiente nos ha caído del cielo. Así, Wickramasinghe no ha dudado en proponer a comienzos de 2020 que la pandemia de COVID-19 se originó con una «lluvia de estrellas» observada desde China en octubre de 2019 (las Oriónidas, fragmentos de la cola del cometa Halley desintegrándose en nuestra atmósfera), que trajo a nuestro planeta el coronavirus SARS-CoV-2. Nada menos. En paralelo, durante la década de 1960 se propuso una variante conocida como «panspermia dirigida», que es aún más imaginativa: la vida terrestre se habría desarrollado a partir de microorganismos formados en otro lugar del Universo, que fueron sembrados deliberadamente aquí por una inteligencia extraterrestre. Aunque algunos investigadores relevantes apoyaron en algún momento esta hipótesis, en la actualidad no tiene ningún sentido hablar sobre ello en un contexto científico.
Más allá de la constatación de que los microorganismos pueden sobrevivir dentro de las rocas, no hay ninguna prueba a favor de cualquiera de las variantes de la panspermia microbiana. Además, en realidad esta hipótesis no responde a las preguntas sobre el origen de la vida: únicamente las cambia de lugar. Por otra parte, si con ella se pretende resolver el problema del tiempo disponible hasta la aparición de los primeros seres vivos, hay que recordar que todo el Sistema Solar (desde donde podrían habernos llegado esos supuestos microorganismos a bordo de meteoritos o núcleos cometarios) se formó a la vez, hace unos 4.570 Ma.
Sin embargo, frente a esta panspermia microbiana altamente improbable, es perfectamente posible que se produjera una «panspermia molecular», es decir, que los mensajeros del espacio llegados masivamente a la Tierra primitiva aportaran una serie de compuestos orgánicos que pudieron ser fundamentales para el origen de la vida terrestre. Haré un símil gastronómico. Imaginemos que en un pueblo de Castilla varios cocineros creativos están preparando una sopa (prebiótica, por supuesto) con una mezcla de los mejores ingredientes que tienen a mano... pero el sabor deseado no acaba de conseguirse: en ese momento llega (como caído del cielo) un chef japonés con un frasco de wasabi, producto desconocido en aquellas tierras, y con él le da el toque exótico que convierte esa sopa en una receta que será recordada durante generaciones.
A LA LUZ DE LA EVOLUCIÓN
Volviendo a la historia de la vida, a partir de un origen terrestre (como pensamos) o extraterrestre (mucho más improbable) de LUCA, la primera bifurcación desde el tronco del árbol originó dos grandes ramas de seres vivos unicelulares, que constituyen los dominios filogenéticos Bacteria y Archaea. Bacterias y arqueas son microorganismos procarióticos (es decir, sin núcleo definido) muy diferentes entre sí, y su progresiva diversificación les permitió colonizar numerosos nichos ecológicos del planeta, desarrollando variados metabolismos. Algunos de ellos dejaron los primeros fósiles de los que tenemos constancia. Aunque continuamente se proponen alternativas, hay un amplio consenso sobre la antigüedad de 3.430 Ma para las primeras huellas fósiles conocidas. Corresponden a «estromatolitos»: capas mineralizadas de comunidades formadas por bacterias (similares a los tapetes microbianos actuales) en las que distintas especies interaccionaban entre sí y establecían relaciones ecológicas.
Mientras tanto, el árbol de la vida seguía creciendo y sus ramas iban divergiendo progresivamente. Pero además de esa diversificación «vertical» existían (y existen) muchas conexiones entre ramas que muestran la transferencia horizontal de genes producida constantemente entre distintos linajes, y a la cual los virus y otras entidades replicativas subcelulares contribuyen decisivamente. Por tanto, aunque al carecer de metabolismo los virus no pueden considerarse auténticos seres vivos, han sido (y seguirán siendo) agentes fundamentales para la evolución de la vida. Si imaginamos los genomas celulares como poemas, los virus serían versos sueltos que entran y salen de ellos. Así, como resultado de esta constante «mezcla» de ramas, se ha propuesto que en lugar de la metáfora del árbol deberíamos imaginar la evolución como un arbusto o, mejor, una enredadera.
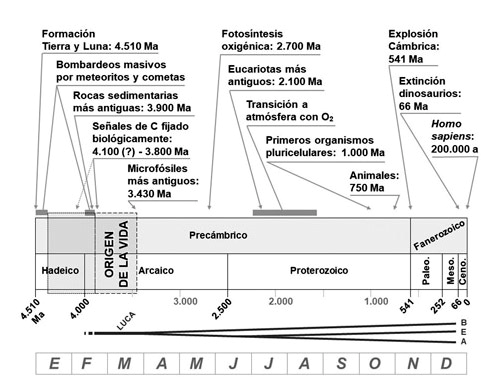
Representación de la historia de la Tierra, dividida en los correspondientes eones y eras, mostrando algunas de las principales etapas en el origen y la evolución de la vida. Bajo la línea de tiempo aparece el árbol que se diversifica hasta formar los tres dominios filogenéticos: Bacteria, Eukarya y Archaea. En la parte inferior se muestra la equivalencia de los 4.510 Ma con un año: las 0:00:00 h del 1 de enero corresponderían al origen del sistema Tierra-Luna, en marzo comenzaría la vida (como la primavera en un año real) y a las 23:59:59 h del 31 de diciembre estaríamos en el momento actual.
A partir de este punto, abrochémonos los cinturones porque vamos a acelerar al máximo la película de la coevolución biosfera-Tierra. La tectónica de placas se instauró de forma global en nuestro planeta hace entre 3.300 y 2.500 Ma, por lo que al comienzo del eón Proterozoico ya se había generalizado el reciclaje de materiales que resulta fundamental para el mantenimiento de la vida. Hace unos 2.700 Ma algunas bacterias acuáticas comenzaron a realizar fotosíntesis oxigénica, un tipo de metabolismo autotrófico que aprovechaba la luz del Sol para fijar CO2, y como subproducto generaba oxígeno molecular (O2). Este era un gas prácticamente inexistente en nuestro planeta, y en primer lugar reaccionó con la gran cantidad de compuestos oxidables que estaban disueltos en los océanos. Finalizado este proceso, el oxígeno que se seguía biosintetizando comenzó a burbujear desde las aguas y a acumularse en la atmósfera hace unos 2.000 Ma. Ante la aparición de ese gas, venenoso para la vida de la época, algunos microorganismos se refugiaron en nichos anóxicos y mantuvieron sus metabolismos anaerobios anteriores, mientras que otros comenzaron a utilizarlo desarrollando una nueva estrategia metabólica, muy eficiente desde el punto de vista energético: la respiración aerobia.
Hace entre 2.000 y 1.500 Ma aparecieron los primeros representantes del dominio filogenético Eukarya, el tercero en el que se clasifican los seres vivos. Las células eucariotas son estructuralmente más complejas que las procariotas, mostrando un núcleo definido que contiene su genoma, así como un complejo sistema de membranas internas y orgánulos especializados. El origen de los primeros microorganismos eucarióticos es un tema de investigación muy activo en la actualidad, y se han planteado diversos modelos basados en la simbiosis entre bacterias y arqueas, en los que estas últimas originaron el núcleo eucariótico. Sin entrar en detalles, una de las hipótesis se basa en la evolución a partir de un tipo concreto de arqueas (pertenecientes al linaje Asgard , cuyo nombre es un guiño a la mitología nórdica), que habrían interiorizado una bacteria (similar a las alfaproteobacterias actuales) de la cual provendrían las mitocondrias, orgánulos clave en el metabolismo energético eucariótico. Un modelo alternativo apunta a la internalización de la arquea dentro del citoplasma de una deltaproteobacteria, seguida de un nuevo evento de endosimbiosis que daría lugar a las mitocondrias.
En cualquier caso, somos el resultado de uno o más procesos de fagocitosis o digestión celular que no llegaron a completarse: es decir, el fruto de la indigestión. Posteriormente, en una rama eucariótica del árbol se produjo otra endosimbiosis, esta vez de cianobacterias, que originó los cloroplastos de los eucariotas fotosintéticos: las algas verdes y rojas, y más tarde las plantas.
QUIÉNES SOMOS
Hace unos 1.000 Ma, la atmósfera terrestre tenía una composición ya muy similar a la actual: 78 % de N2 , 21 % de O2, 0,9 % de argón y trazas de CO2 y otros gases. Durante esa época, gracias entre otros factores al O2 disponible, en diferentes linajes de eucariotas apareció una forma de organización que las bacterias apenas habían explorado: la pluricelularidad. De los cuatro reinos eucarióticos que distinguimos en la actualidad, los protistas siguen siendo unicelulares mientras que los otros tres (plantas, hongos y animales) son pluricelulares. La vida pluricelular es la única que vemos sin ayuda del microscopio (y por tanto la habitualmente considerada cuando se habla de preservar la biodiversidad) pero representa sólo el 15 % de las especies conocidas.
Realmente la Tierra es, desde su origen, un planeta dominado por los microorganismos. En cualquier otro lugar donde se pueda haber originado la vida, aunque fuera bioquímicamente similar a la nuestra, su evolución no tendría por qué haber conducido a la aparición de los eucariotas... y mucho menos a los pluricelulares. Convertirse en un organismo pluricelular requiere que las células (inicialmente iguales entre sí) se vayan especializando en realizar distintas funciones. En algunos linajes eucarióticos esto condujo a un sistema de reproducción sexual, muy ventajoso para generar diversidad, en el que las células germinales de los progenitores pervivirán en la siguiente generación, mientras que todas las demás (llamadas somáticas) dejan de ser necesarias una vez que el proceso reproductivo ha terminado. Con ello aparecía en nuestro planeta algo que no era una consecuencia de la vida (iniciada 2.500 Ma antes) sino de la pluricelularidad y la reproducción sexual: la muerte.
La evolución posterior de los organismos pluricelulares estuvo condicionada por la Explosión cámbrica, una etapa de diversificación rápida iniciada hace 540 Ma que condujo a la aparición de los planes anatómicos de la mayor parte de los animales actuales, y a la diferenciación clara entre depredadores y presas. Hace unos 66 Ma, la caída de un gran meteorito produjo la extinción masiva de biodiversidad ocurrida a finales del Cretácico, en la que desapareció buena parte de las especies pluricelulares incluyendo la totalidad de los dinosaurios. Así, aquel impacto (de nuevo, un evento casual) permitió que otros grupos de animales, en particular los mamíferos, se diversificaran y ocuparan numerosos nichos ecológicos. Actualmente, en el último fotograma de esta larga y trepidante película, nuestro planeta acoge cientos de millones de especies unicelulares y pluricelulares, junto a innumerables virus y otras entidades replicativas que las parasitan, conectadas entre sí a través de complejas redes ecológicas.
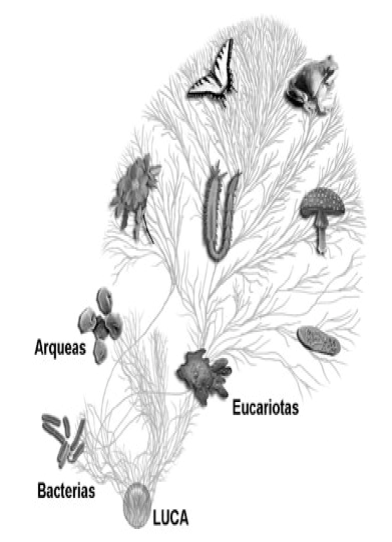
Representación esquemática del árbol de la vida, mostrando los tres dominios filogenéticos (con un sesgo claro hacia los eucariotas pluricelulares, que sólo deberían suponer el 15 % del total) y algunas ramas que los conectan mediante transferencia horizontal de genes
Y, entre toda esta biodiversidad, nosotros. Desde el punto de vista biológico, los humanos no somos especiales, ni éramos necesarios, ni había nada en la Química de aquella sopa primitiva que permitiera predecir nuestro origen... o que «tendiera» hacia él. No ocupamos la cima de una inexistente pirámide evolutiva, sino una más de las ramas que constituyen el árbol de la vida. Otro fruto del azar y la necesidad. Pero los paleoantropólogos nos enseñan que, a lo largo del linaje del género Homo, las capacidades de nuestro cerebro se han ido desarrollando de forma imparable y nos han llevado a protagonizar una rápida evolución cultural. Hemos transformado el mundo, escribimos poemas y miramos al cielo preguntándonos si estamos solos.
Mientras tanto, recordando a Galileo Galilei, «el Sol, con todos los planetas que rotan a su alrededor y dependen de él, sigue haciendo madurar los racimos de uvas como si no tuviese otra cosa que hacer en el Universo».
© 2023 JAVIER DE LUCAS